To support its specialized functions, the skin has basic requirements that must be satisfied for almost every tissue. It needs mechanical strength, largely provided by a supporting framework of extracellular matrix, mainly secreted by fibroblasts. It needs a blood supply to bring nutrients and oxygen and remove waste products and carbon dioxide, and this requires a network of blood vessels, lined with endothelial cells. These vessels also provide access routes for cells of the immune system to provide defenses against infection: macrophages and dendritic cells phagocytose invading pathogens and help activate lymphocytes, which mediate more sophisticated adaptive immune system responses (discussed in Chapter 24). Nerve fibers are needed too, to convey sensory information from the tissue to the central nervous system, and to deliver signals in the opposite direction for glandular secretion and smooth muscle contraction.
illustrates the architecture of the tissue and shows how it makes provision for all these support services. Skin consists of two main parts: an epithelium, the epidermis, lying outermost, and beneath this a layer of connective tissue, which includes the tough collagen-rich dermis (from which leather is made) and the underlying fatty subcutaneous layer or hypodermis. In the skin, as elsewhere, the connective tissue, with vessels and nerves running through it, is responsible for most of the general supportive functions listed above.
Mammalian skin. (A) These diagrams show the cellular architecture of thick skin. (B) Micrograph of a cross section through the sole of a human foot, stained with hematoxylin and eosin. The skin can be viewed as a large organ composed of two main tissues: (more...)
The defining component of the skin—the specialized tissue that is peculiar to this organ, even though not the major part of its bulk—is the epidermis. This has a simple organization, and it provides a beautiful introduction to the way in which tissues of the adult body are continually renewed, through processes similar to those that operate in the embryo. We return to connective tissues later.
Epidermal Cells Form a Multilayered Waterproof Barrier
The epidermis suffers more direct, frequent, and damaging encounters with the external world than any other tissue in the body. Its need for repair and renewal is central to its organization.
The epidermis is a multilayered (stratified) epithelium composed largely of keratinocytes (so named because their characteristic differentiated activity is the synthesis of intermediate filament proteins called keratins, which give the epidermis its toughness) (). These cells change their appearance from one layer to the next. Those in the innermost layer, attached to an underlying basal lamina, are termed basal cells, and it is usually only these that divide. Above the basal cells are several layers of larger prickle cells (), whose numerous desmosomes—each a site of anchorage for thick tufts of keratin filaments—are just visible in the light microscope as tiny prickles around the cell surface (hence the name). Beyond the prickle cells lies the thin, darkly staining granular cell layer (see ). It is at this level that the cells are sealed together to form a waterproof barrier, fulfilling the most fundamentally important of all the functions of the epidermis. Mice that fail to form this barrier because of a genetic defect die from rapid fluid loss soon after birth, even though their skin appears normal in other respects.
The multilayered structure of the epidermis, as seen in a mouse. The outlines of the keratinized squames are revealed by swelling them in a solution containing sodium hydroxide. The highly ordered hexagonal arrangement of interlocking columns of cells (more...)
A prickle cell. This drawing, from an electron micrograph of a section of the epidermis, shows the bundles of keratin filaments that traverse the cytoplasm and are inserted at the desmosome junctions that bind the prickle cell (red) to its neighbors. (more...)
The granular layer, with its barrier to the movement of water and solutes, marks the boundary between the inner, metabolically active strata and the outermost layer of the epidermis, consisting of dead cells whose intracellular organelles have disappeared. These outermost cells are reduced to flattened scales, or squames, filled with densely packed keratin. The plasma membranes of both the squames and the outer granular cells are reinforced on their cytoplasmic surface by a thin (12 nm), tough, cross-linked layer of proteins, including a cytoplasmic protein called involucrin. The squames themselves are normally so compressed and thin that their boundaries are hard to make out in the light microscope, but soaking in sodium hydroxide solution (or a warm bath tub) makes them swell slightly, and their outlines can then be seen (see ).
Differentiating Epidermal Cells Synthesize a Sequence of Different Keratins as They Mature
Having described the static picture, let us now set it in motion and see how the epidermis is continually renewed by the production of new cells in the basal layer. While some basal cells are dividing, adding to the population in the basal layer, others (their sisters or cousins) are slipping out of the basal cell layer into the prickle cell layer, taking the first step on their outward journey. When they reach the granular layer, the cells start to lose their nucleus and cytoplasmic organelles, through a degradative mechanism that involves partial activation of the machinery of apoptosis; in this way, the cells are transformed into the keratinized squames of the keratinized layer. These finally flake off from the surface of the skin (and become a main constituent of household dust). The period from the time a cell is born in the basal layer of the human skin to the time it is shed from the surface is of the order of a month, depending on the region of the body.
The accompanying molecular transformations can be studied by analyzing either thin slices of epidermis cut parallel to the surface or successive layers of cells stripped off by repeatedly applying and removing strips of adhesive tape. The keratin molecules, for example, which are plentiful in all layers of the epidermis, can be extracted and characterized. They are of many types (discussed in Chapter 16), encoded by a large family of homologous genes, with the variety further increased through alternative RNA splicing. As the new keratinocyte in the basal layer is transformed into the squame in the outermost layers (see ), it switches from one selection of keratins to another. Meanwhile other characteristic proteins, such as involucrin, also begin to be synthesized as part of a coordinated program of terminal cell differentiation—the process in which a precursor cell acquires its final specialized characteristics and usually permanently stops dividing. The whole program is initiated in the basal layer. It is here that the fates of the cells are decided.
Epidermis Is Renewed by Stem Cells Lying in Its Basal Layer
The outer layers of the epidermis are replaced a thousand times over in the course of a human lifetime. In the basal layer there have to be cells that can remain undifferentiated and carry on dividing for this whole period, continually throwing off descendants that differentiate, leave the basal layer, and are eventually discarded. The process can be maintained only if the basal cell population is self-renewing. It must therefore contain some cells that generate a mixture of progeny, including daughters that remain undifferentiated like their parent, as well as daughters that differentiate. Cells with this property are called stem cells. They have so important a role in such a variety of tissues that it is useful to have a formal definition.
The defining properties of a stem cell are as follows:
- 1.
It is not itself terminally differentiated (that is, it is not at the end of a pathway of differentiation).
- 2.
It can divide without limit (or at least for the lifetime of the animal).
- 3.
When it divides, each daughter has a choice: it can either remain a stem cell, or it can embark on a course that commits it to terminal differentiation ().
The definition of a stem cell. Each daughter produced when a stem cell divides can either remain a stem cell or go on to become terminally differentiated. In many cases, the daughter that opts for terminal differentiation undergoes additional cell divisions (more...)
Although it is part of the definition of a stem cell that it should be able to divide, it is not part of the definition that it should divide rapidly; in fact, stem cells usually divide at a relatively low rate. They are required wherever there is a recurring need to replace differentiated cells that cannot themselves divide, and this includes a great variety of tissues. Thus stem cells are of many types, specialized for the genesis of different classes of terminally differentiated cells—epidermal stem cells for epidermis, intestinal stem cells for intestinal epithelium, hemopoietic stem cells for blood, and so on. Each stem-cell system nevertheless raises similar fundamental questions. What factors determine whether the stem cell divides or stays quiescent? What decides whether a given daughter cell differentiates or remains a stem cell? And where the stem cell can give rise to more than one kind of differentiated cell—as is very often the case—what determines which differentiation pathway is followed?
The Two Daughters of a Stem Cell Do Not Always Have to Become Different
At steady state, to maintain a stable stem-cell population, precisely 50% of the daughters of stem cells in each cell generation must remain as stem cells. In principle, this could be achieved in two ways—through environmental asymmetry or through divisional asymmetry (). In the one strategy, the division of a stem cell could generate two initially similar daughters whose fates would be governed by their subsequent environment; 50% of the population of daughters would remain as stem cells, but the two daughters of an individual stem cell in the population might often have the same fate. At the opposite extreme, the stem cell division could be always strictly asymmetric, producing one daughter that inherits the stem-cell character and another that inherits factors that force it to embark on differentiation. In the latter case, the existing stem cells could never increase their numbers, and any loss of stem cells would be irreparable.
Two ways for a stem cell to produce daughters with different fates. In the strategy based on environmental asymmetry, the daughters of the stem cell are initially similar and are directed into different pathways according to the environmental influences (more...)
In fact, if a patch of epidermis is destroyed, the damage is repaired by surrounding epidermal cells that migrate in and proliferate to cover the denuded area. In this process, a new self-renewing patch of epidermis is established, implying that additional stem cells have been generated to make up for the loss. These must have been produced by symmetric divisions in which one stem cell gives rise to two. In this way, the stem cell population adjusts its numbers to fit the available niche.
Observations such as these suggest that the maintenance of stem cell character in the epidermis might be controlled by contact with the basal lamina, with a loss of contact triggering the start of terminal differentiation, and maintenance of contact tending to preserve stem cell potential. This idea contains a grain of truth, but it is not the whole truth, as we now explain.
The Basal Layer Contains Both Stem Cells and Transit Amplifying Cells
Basal keratinocytes can be dissociated from intact epidermis and can proliferate in a culture dish, giving rise to new basal cells and to terminally differentiated cells. Even within a population of cultured basal keratinocytes that all seem undifferentiated, there is great variation in the ability to proliferate. When cells are taken singly and tested for their ability to found new colonies, some seem unable to divide at all, others go through only a few division cycles and then halt, and still others divide enough times to form large colonies. This proliferative potential directly correlates with the expression of the β1 subunit of integrin (see ). Clusters of cells with high levels of this molecule can be found in the basal layer of the intact human epidermis also, and they are thought to be the stem cells ().
The distribution of stem cells in human epidermis, and the pattern of epidermal cell production. The diagram is based on specimens in which the location of the stem cells was identified by staining for β1-integrin, and that of the differentiating (more...)
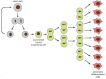
Basal cells expressing β1 integrin at a lower level can also divide—indeed, they divide more frequently—but only for a limited number of division cycles, after which they leave the basal layer and differentiate. These latter cells are called transit amplifying cells—“transit”, because they are in transit from a stem-cell character to a differentiated character; “amplifying”, because the division cycles they go through have the effect of amplifying the number of differentiated progeny that result from a single stem-cell division (). Mingled with the population of transit amplifying cells are some cells, still connected with the basal lamina by a thin stalk, that have already stopped dividing and begun to differentiate, as indicated by the types of keratin molecules they express. Contact with the basal lamina, therefore, cannot be the only factor controlling the developmental fate of an epidermal basal cell.
Transit amplifying cells. Stem cells in many tissues divide only rarely, but give rise to transit amplifying cells—daughters committed to differentiation that go through a limited series of more rapid divisions before completing the process. In (more...)
This is not to say that contact with the basal lamina or a similar substratum does not matter. If cultured basal keratinocytes are held in suspension, instead of being allowed to settle and attach to the bottom of the culture dish, they all stop dividing and differentiate. To remain as an epidermal stem cell, it is apparently necessary for it to be attached to the basal lamina or other extracellular matrix, even though it is not sufficient. This requirement helps ensure that the size of the stem cell population does not increase without limit. If crowded out of their regular niche on the basal lamina, the cells lose their stem cell character. When this rule is broken, as in some cancers, the result can be an ever-growing tumor.
Epidermal Renewal Is Governed by Many Interacting Signals
Cell turnover in the epidermis seems at first glance a simple matter, but the simplicity, as we have just seen, is deceptive. There are many points in the process that have to be controlled according to circumstances: the rate of stem-cell division; the probability that a stem-cell daughter will remain a stem cell; the number of cell divisions of the transit amplifying cells; the timing of exit from the basal layer, and the time that the cell then takes to complete its differentiation program and be sloughed from the surface. Regulation of these steps must enable the epidermis to respond to rough usage by becoming thick and callused, and to repair itself when wounded. In specialized regions of epidermis, such as those that form hair follicles, with their own specialized subtypes of stem cells, yet more controls are needed to organize the local pattern.
Each control point is important in its own way, and a whole panoply of molecular signals regulate them, so as to keep the body surface always properly covered. Most of the cell communication mechanisms described in Chapter 15 are implicated, either in signaling between cells within the epidermis or in signaling between epidermis and dermis. The EGF, FGF, Wnt, Hedgehog, Notch, BMP/TGFβ, and integrin signaling pathways are all involved (and we shall see that the same is true of most other tissues). Mutations in components of the Hedgehog or Wnt pathways, for example, can lead to the development of epidermal cancers. Components of the Hedgehog, Notch, BMP, and Wnt pathways when misexpressed interfere with the formation of hairs, blocking their development or causing them to develop out of place.
Activation of the Wnt pathway seems to favor maintenance of stem-cell character, inhibiting the switch from stem cell to transit amplifying cell, whereas Notch signaling in the epidermis seems to have a contrary effect, inhibiting neighbors of stem cells from remaining as stem cells. TGFβ has a key role in signaling to the dermis during the repair of skin wounds, promoting the formation of collagen-rich scar tissue. And integrins in the epidermis are not merely markers of cell character, but also regulators of cell fate. Thus, when transgenic mice are engineered to maintain in upper epidermal layers the expression of integrins normally confined to the basal layer, they develop a condition resembling the common human skin disorder psoriasis: the rate of basal cell proliferation is greatly increased, the epidermis thickens, and cells are shed from the surface of the skin within as little as a week after emerging from the basal layer, before they have had time to keratinize fully. The precise individual functions of all the various signaling mechanisms in the epidermis are only beginning to be disentangled.
The Mammary Gland Undergoes Cycles of Development and Regression
In specialized regions of the body surface, other types of cells besides the keratinized cells described above develop from the embryonic epidermis. In particular, secretions such as sweat, tears, saliva, and milk are produced by cells segregated in deep-lying glands that originate as ingrowths of the epidermis. These epithelial structures have functions and patterns of renewal quite different from those of keratinizing regions.
The mammary glands are the largest and most remarkable of these secretory organs. They are the defining feature of mammals and an important concern in many ways: not only for nourishment of babies and attraction of the opposite sex, but also as the basis for a large industry—the dairy industry—and as the site of some of the commonest forms of cancer. Mammary tissue illustrates most dramatically that developmental processes continue in the adult body; and it shows how cell death by apoptosis can allow development to go into reverse.
Milk production must be switched on when a baby is born and switched off when the baby is weaned. A “resting” adult mammary gland consists of branching systems of ducts embedded in fatty connective tissue. The ducts are lined by an epithelium that includes a subpopulation of mammary stem cells (though the precise location of the stem cells is still debated). As a first step toward milk production, the hormones that circulate during pregnancy cause the duct cells to proliferate, increasing their numbers tenfold or twentyfold. The terminal portions of the ducts grow and branch, forming little dilated outpocketings, or alveoli, containing secretory cells (). Milk secretion begins only when these cells are stimulated by the different combination of hormones that circulate in the mother after the birth of the baby. A further tier of hormonal control governs the actual ejection of milk from the breast: the stimulus of suckling causes cells of the hypothalamus (in the brain) to release the hormone oxytocin, which travels via the bloodstream to act on myoepithelial cells. These musclelike cells originate from the same epithelial precursor population as the secretory cells of the breast, and they have long spidery processes that embrace the alveoli. In response to oxytocin they contract, thereby squirting milk out of the alveoli into the ducts.
The mammary gland. (A) The growth of alveoli from the ducts of the mammary gland during pregnancy and lactation. Only a small part of the gland is shown. The “resting” gland contains a small amount of inactive glandular tissue embedded (more...)
Eventually, when the baby is weaned and suckling stops, the secretory cells die by apoptosis, and most of the alveoli disappear. Macrophages rapidly clear away the dead cells, and the gland reverts to its resting state. This ending of lactation is abrupt and, unlike the events that lead up to it, seems to be induced by the accumulation of milk, rather than by a hormonal mechanism. If one subset of mammary ducts is obstructed so that no milk can be discharged, the secretory cells that supply it commit mass suicide by apoptosis, while other regions of the gland survive and continue to function. The apoptosis is triggered by a combination of factors including TGFβ3, which accumulates where milk secretion is blocked ().
Death of milk-secreting cells when suckling stops. (A) A section of part of a lactating mammary gland of a mouse that had been suckling her newborn babies in the normal way. (B) A corresponding section from a mouse that had not suckled for 9 hours. The (more...)
Cell division in the growing mammary gland is regulated not only by hormones but also by local signals passing between cells within the epithelium and between the epithelial cells and the connective tissue, or stroma, in which the epithelial cells are embedded. All the signals listed earlier as important in controlling cell turnover in the epidermis are also implicated in controlling events in the mammary gland. Again, signals delivered via integrins play a crucial part: deprived of the basal lamina adhesions that activate integrin signaling, the epithelial cells fail to respond normally to hormonal signals. Faults in these interacting control systems underlie some of the commonest forms of cancer, and we need to understand them better.
Summary
Skin consists of a tough connective tissue, the dermis, overlaid by a multilayered waterproof epithelium, the epidermis. The epidermis is continually renewed from stem cells, with a turnover time, in humans, of the order of a month. Stem cells, by definition, are not terminally differentiated and have the ability to divide throughout the lifetime of the organism, yielding some progeny that differentiate and others that remain stem cells. The epidermal stem cells lie in the basal layer, attached to the basal lamina. Progeny that become committed to differentiation go through several rapid divisions in the basal layer, and then stop dividing and move out toward the surface of the skin. They progressively differentiate, switching from expression of one set of keratins to expression of another until, eventually, their nuclei degenerate, producing an outer layer of dead keratinized cells that are continually shed from the surface.
The fate of the daughters of a stem cell is controlled in part by interactions with the basal lamina and in part by a variety of signals from neighboring cells. These controls allow two stem cells to be generated from one during repair processes, and they regulate the rate of basal cell proliferation according to need. Glands connected to the epidermis, such as the mammary glands, have their own stem cells and their own distinct patterns of cell turnover, governed by other conditions. In the breast, for example, circulating hormones stimulate the cells to proliferate, differentiate, and make milk; the cessation of suckling triggers the milk-secreting cells to die by apoptosis, as a result of a build-up of TGFβ3 where milk fails to be drained away.