

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Ebert D. Ecology, Epidemiology, and Evolution of Parasitism in Daphnia [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2005.

This chapter provides an overview of the biology of waterfleas of the genus Daphnia. It describes basic aspects of individual physiology and nutrition, including some remarks about immunity. It summarizes the typical life cycle and development of Daphnia. The modes of reproduction and the induction of resting egg production in cyclic and obligate parthenogenetic forms are discussed. Finally, population biological aspects, such as behavioral ecology, habitat preferences, population genetics, and population dynamics, are introduced.
Introduction
The following introduction summarizes the key aspects of the natural history of the genus Daphnia. It is far from exhaustive. Because much of the biology has been studied over the past 250 years and is considered to be common knowledge, I give no references to specific studies in most cases. The interested reader may consult Freyer (1991), Kästner (1993), Lampert and Sommer (1999), and Peters and De Bernardi (1987), from which much of the information was taken. A search on the Internet using the term "Daphnia" or "Cladocera" will also bring up a large body of information. Of particular interest is the Cladocera Web page of the University of Guelph in Canada. The reader familiar with Daphnia may skip this chapter and move on to Chapter 3.
Daphnia are planktonic crustaceans that belong to the Phyllopoda (sometimes called Branchiopoda), which are characterized by flattened leaf-like legs used to produce a water current for the filtering apparatus. Within the branchiopods, Daphnia belong to the Cladocera, whose bodies are enclosed by an uncalcified shell (Figures 2.1 and 2.2), known as the carapace. It has a double wall, between which hemolymph flows and which is part of the body cavity. The carapace is largely made of chitin, a polysaccharide. Cladocera have up to 10 pairs of appendages, which are (from front to back): antennules, antennae (the second antennae, used for swimming); maxillae; and mandibles; followed by 5 (as in Daphnia) or 6 limbs on the trunk. The limbs form an apparatus for feeding and respiration. At the end of the abdomen is a pair of claws. The body length of Cladocera ranges from less than 0.5 mm to more than 6 mm. Males are distinguished from females by their smaller size, larger antennules, modified post-abdomen, and first legs, which are armed with a hook used in clasping.

Figure 2.1
The functional anatomy of Daphnia .

Figure 2.2
Ventral view of Daphnia Adult females. On the left, a photograph of Daphnia longispina from a rock pool population in southern Finland. On the right, a drawing. (The drawing is reproduced with permission from Kästner: Lehrbuch der speziellen Zoologie, (more...)
The genus Daphnia includes more than 100 known species of freshwater plankton organisms found around the world (see Figures 2.3, 2.4, and 2.5 for three European representatives of the genus). They inhabit most types of standing freshwater except for extreme habitats, such as hot springs. All age classes are good swimmers and are mostly pelagic, i.e., found in the open water. They live as filter feeders, but some species may frequently be seen clinging to substrates such as water plants or even browsing over the bottom sediments of shallow ponds. Adults range from less than 1 mm to 5 mm in size, with the smaller species typically found in ponds or lakes with fish predation. The ecology of the genus Daphnia may be better known than the ecology of any other group of organisms.

Figure 2.3
Daphnia magna . Adult female with a clutch of freshly laid parthenogenetic eggs in its brood chamber. The female is from a laboratory culture of a clone originating in a pond near Oxford, UK.
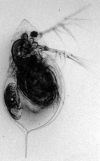
Figure 2.4
Daphnia cucullata . Adult female with one embryo in her brood chamber. This female comes from a laboratory culture of a clone originally isolated from Klostersee in southern Bavaria, Germany.

Figure 2.5
Daphnia longispina . Adult female with three eggs in the brood chamber. This female was isolated from a rock pool in southern Finland close to Tvärminne.
Physiology, Metabolism, and Immunity
Daphnia feed on small, suspended particles in the water. They are suspension feeders (filter feeders). The food is gathered with the help of a filtering apparatus, consisting of the phylopods, which are flattened leaf-like legs that produce a water current. As the current flows anterior to posterior, the Daphnia collect particles that are transferred into the food groove by special setae. Although the feeding apparatus is so efficient that even bacteria can be collected, the food is usually made up of planktonic algae. Green algae are among the best food, and most laboratory experiments are done with either Scenedesmus or Chlamydomonas, both of which are easy to culture in monoclonal chemostats. Daphnia usually consume particles from around 1 μm up to 50 μm, although particles of up to 70 μm in diameter may be found in the gut content of large individuals.
The dynamics of food uptake follow a functional response type 1. Below a certain food concentration (the incipient limiting level), the food uptake from the water (feeding rate) is proportional to the food concentration, and the filtering rate (amount of water filtered per unit time) is maximal. Above this level, the feeding rate is constant because the filtering rate decreases with increasing food concentration in the water. For parasites that enter the host with the food particles, infection rates depend on the food concentration in the water. Highest infection rates are expected when filtering rates are maximal.
The gut is more or less tubular with three parts: the esophagus, the midgut, and the hindgut. There are two small digestive ceca (diverticula) that are easily seen in the head section of the midgut (Figures 2.1 and 2.6). The midgut is lined with an epithelium and bears microvilli. Peristaltic contractions of the gut wall pass food through the gut, but a peritrophic membrane contains the food and prevents it from entering the ceca. Epithelial cells do not phagocytose particles but absorb molecules. The pH is 6 to 6.8 in the anterior part of the midgut and 6.6 to 7.2 in the posterior part. Food is expelled from the hindgut by peristaltic movement but also requires the pressure of more recently acquired food particles. The color of Daphnia adapts to the food that is predominant in their diet. Daphnia feeding on green algae will be transparent with a tint of green or yellow, whereas those feeding on bacteria will be white or salmon-pink. Well-fed animals are more strongly colored than starved animals.

Figure 2.6
Gut of Daphnia magna . Gut dissected from a female. On the left, the paired intestinal ceca can be seen. The gut ends at the right side. The esophagus cannot be seen in this preparation. The dark material is partially digested gut content.
Daphnia have an open blood circulation. The heart is located dorsally and anterior from the brood chamber. At 20ºC, it beats about 200 times per minute, slowing down at lower temperatures. Blood cells are easily visible through the transparent body as they flow rapidly through the body cavity. To support oxygen transport, Daphnia have the extracellular respiratory protein hemoglobin (Hb), a multi-subunit, multi-domain macromolecule. There are at least four Hb genes. Daphnia tend to develop more Hb to increase oxygen uptake from the water. In response to environmental changes (oxygen concentration, temperature), the Hb concentration varies up to about 20-fold. Oxy-hemoglobin, the form that is loaded with oxygen, is red and gives the transparent animals a reddish appearance (Figure 2.7). Because certain parasites also cause the hemolymph to become red, one cannot easily determine the cause of the red color from sight alone. However, low oxygen usually affects an entire population, coloring all animals reddish, whereas parasites usually infect only a portion of the population.
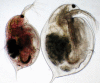
Figure 2.7
Two Daphnia magna with contrasting hemolymph color due to haemoglobin (Hb). These two adult females were taken from two adjacent rock pool populations in southern Finland at a time when one of the pools was low in oxygen (because of overfertilization (more...)
Daphnia have the usual osmoregulatory problems of freshwater animals, i.e., too much water and too few solutes. They are able to absorb ions with chloride-absorbing glands. The shell gland (maxillary gland; Figure 2.1) may have a role in excretion and/or osmoregulation.
The nervous system is characterized by the cerebral ganglion, which is located close to the gut and near the eye. Juvenile and adult Daphnia have one large compound eye, whereas embryos show two brownish eye spots that fuse during the last part of the development. The compound eye helps to orient the animal while swimming. A small structure called a naupliar eye is located between the mouth and the compound eye on top of the cerebral ganglion.
Many invertebrates, including crustaceans, have a well-developed innate immune system that includes melanization by activation of the prophenoloxidase (proPO) activating system, a clotting process, phagocytosis, encapsulation of foreign material, antimicrobial action, and cell agglutination (Söderhall 1999). Of these mechanisms, only the proPO system (Mucklow and Ebert 2003) and phagocytosis (Metchnikoff 1884) have been identified in Daphnia, although there is no reason to doubt that the other mechanisms work as well. The proPO system is not only a defense system against parasites but is also involved in wound healing of the cuticle (Figure 2.8). The enzyme involved in melanin formation, PO, has been detected in the blood of many arthropods, including D. magna. Melanin is a brown pigment that is also used for carapace pigmentation in some Daphnia, particularly Daphnia in the high Arctic, where the dark pigment protects them from uninterrupted solar radiation (repair of UV damage is only possible in the dark). Acquired immunity is thought to be absent in invertebrates; however, transmission of strain-specific immunity from mothers to offspring has recently been suggested for D. magna (Little et al. 2003).

Figure 2.8
Wound healing in Daphnia magna . An injury in the carapace heals within a few hours to days. Wound healing involves a melanization reaction that stains the wound dark. Here a needle was used to injure the carapace. The squared pattern in the background (more...)
Life Cycle and Development
The life cycle of Daphnia during the growth season is characterized by its asexual mode of reproduction (apomixis) (Figure 2.9). A female produces a clutch of parthenogenetic (amictic) eggs after every adult molt (if feeding conditions permit). Figures 2.3 to 2.5 show females with parthenogenetic eggs. The eggs are placed in the brood chamber, which is located dorsally beneath the carapace and which is closed by the abdominal processes (Figures 2.1 and 2.10). Development of eggs is direct (immediate). At 20ºC, the embryos hatch from the eggs after about 1 day but remain in the brood chamber for further development (Figure 2.11). After about 3 days in the brood chamber, the young Daphnia are released by the mother through ventral flexion of the post-abdomen. The newborn look more or less like the adult Daphnia, except that the brood chamber is not yet developed (Figure 2.12). In most species, a juvenile Daphnia passes through four to six juvenile instars before it becomes primipare, i.e., produces eggs for the first time. The age at which the first eggs are deposited into the brood chamber is around 5-10 days at 20ºC, but this may take longer under poor feeding conditions. An adult female may produce a clutch of eggs every 3 to 4 days until her death. In the laboratory, females may live for more than 2 months, with a higher age being reached under poorer feeding conditions. Clutch sizes vary among species, from 1 to 2 eggs in small species such as D. cucullata (Figure 2.4) to more than 100 in large species such as D. magna (Figure 2.3).
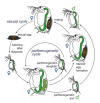
Figure 2.9
Life cycle of a cyclic parthenogenetic Daphnia . This diagram depicts the sexual and the asexual (parthenogenetic) life cycle of a Daphnia. During the parthenogenetic cycle, females produce diploid eggs that develop directly into daughters. The same female (more...)
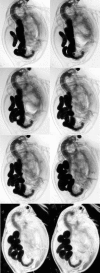
Figure 2.10
Egg laying by Daphnia magna . This series of pictures, taken within a 15-minute time span, shows the process of placing eggs into the brood chamber. The eggs are initially sausage shaped but quickly resume a nearly spherical shape. The ovaries shrink (more...)
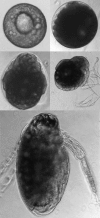
Figure 2.11
Development of Daphnia embryos. This sequence shows the development of parthenogenetic eggs and embryos raised in a culture dish. The top left picture shows an egg within the first 8 hours after egg laying. The last picture shows an embryo of about 2 (more...)

Figure 2.12
Newborn Daphnia magna . A parthenogenetic offspring of D. magna within the first 12 hours after release from the brood chamber.
Although in a typical growth season Daphnia produce diploid (2N) eggs that develop directly and without a resting phase, a different type of egg is produced for resting (Figure 2.9). These resting eggs are encapsulated in a protective, saddle-like structure called an ephippium (Figures 2.13 and 2.14), which is usually strongly melanized and contains 2 large eggs, 1 from each ovary. It is not uncommon, however, to find ephippia with only 1 egg, or none at all. The ephippium is cast off at the next molt. In most cases, these eggs are produced sexually, but obligate parthenogenetic Daphnia, which are typically found in the northern parts of America and Eurasia and are an excellent example of geographic parthenogenesis (best described for D. pulex), may produce diploid resting eggs asexually as well. In the more common sexual Daphnia (with cyclic parthenogenesis), resting egg production follows the asexual production of diploid males, which are needed to fertilize the haploid eggs (Figures 2.15 and 2.16). A sexual clone can produce males and females and is capable of self-fertilization. For fertilization, males copulate with females (sometimes two males can be observed on the same female). Fertilization is internal and happens between molting and deposition of eggs into the ephippium. Sperm are either tailless (Figure 2.17) or may contain short, pseudopodia-like extensions. The induction of sexuality seems to be triggered by a complex set of stimuli, the most important possibly being those that go hand-in-hand with a high Daphnia density, e.g., increased competition and reduced food availability. Abiotic factors alone, such as decreased day length and lowered temperature, also seem to play a minor role. Daphnia in intermittent populations (such as ponds that are likely to dry up during part of the year) have a much higher tendency to produce resting eggs than Daphnia in permanent populations, e.g., in large lakes.

Figure 2.13
Daphnia magna carrying a resting egg. The ephippium is easily recognized by its dark color. It usually contains two eggs, which are haploid and require fertilization. In the central part of the body of this female, the filled, greenish ovaries can be (more...)

Figure 2.14
Daphnia longispina carrying a resting egg. The ephippium is not yet fully developed. Its wall is still transparent, and the two eggs are visible. These eggs are haploid and require fertilization. This female was isolated from a rock pool population (more...)

Figure 2.15
Male Daphnia magna . A male D. magna from a laboratory population of a clone isolated from a rock pool in southern Finland. Photo by Dita B. Vizoso, Fribourg University.
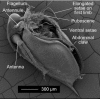
Figure 2.16
Scanning electron microscopic of an adult male Daphnia magna . A male D. magna from a laboratory population of a clone isolated from a rock pool in southern Finland. Photo by Frida Ben-Ami, Basel University.
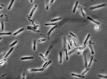
Figure 2.17
Sperm of Daphnia magna . Sperm in D. magna are tailless, rod-shaped, and up to 9 μm in length. Other Daphnia species have sperm with short pseudopodia (not shown here). Sperm is haploid.
The ephippia from the females are released with the shed carapace during molting and sink to the bottom or float with the help of small gas chambers. They may disperse with the wind or with animals (e.g., attached to the feathers of waterfowl) or may drift with the water. Floating ephippia may be driven by winds to form large piles of resting eggs in wind-protected parts of ponds and lakes. Depending on the habitat, resting eggs may endure unfavorable seasons (e.g., winter colds, periods of low bio-productivity, summer droughts), and hatching is induced by external stimuli, such as an appropriate photoperiod, light, rising temperatures, or simply the presence of water in a previously dry pond. From resting eggs, only females hatch, which usually produce parthenogenetic eggs themselves but may directly produce resting stages under conditions of a very short growing season.
Habitat
Daphnia populations can be found in a range of water bodies, from huge lakes down to very small temporary pools, such as rock pools (Figures 2.18 and 2.19) and vernal pools (seasonally flooded depressions). Often they are the dominant zooplanktor and form, as such, an essential part of the food web in lakes and ponds. In many lakes, Daphnia are the predominant food for planktivorous fish, at least at times. As a consequence, the Daphnia species distribution and life history are closely linked with the occurrence of predators. Typically, Daphnia species found in lakes with planktivorous fish are smaller and more transparent than species found in fishless water bodies. Large species such as D. magna and D. pulex usually cannot survive under intensive fish predation, whereas small species such as D. galeata, D. cucullata, and D. hyalina are usually not found in fishless water bodies. A number of invertebrates are known to prey on Daphnia, the best investigated probably being the larvae of the phantom midge Chaoborus and the water boatman Notonecta and related genera. Although visually-hunting fish usually show a preference for larger prey items, invertebrates may prefer smaller prey or even a specific size class. These differences in size-specific mortality rates are believed to be a key factor in the evolution of Daphnia body size. High juvenile mortality caused by Chaoborus has been suggested as the cause for evolution of larger sizes at birth and to phenotypically plastic adjustments of birth size and growth rates. In contrast, predation by fish has led to smaller sizes and earlier maturation age.

Figure 2.18
Rock pools with Daphnia populations on four islands in the Tvärminne archipelago in southern Finland. These pools are part of a metapopulation of D. magna, D. pulex, and D. longispina. In the background, the Baltic Sea is visible.

Figure 2.19
Dry rock pool with Daphnia magna populations on an island in the Tvärminne archipelago in southern Finland. The upper photo shows a dry pool in July 2003. The lower photo shows a close-up of the sediment surface in the pool. The dry sediment has (more...)
It has been proposed that in water bodies without predation, the composition of Daphnia species is influenced by size-dependent competition, with larger species out-competing smaller species. There is, however, some debate about the efficiency of size-dependent processes in the absence of predation.
The water quality of Daphnia habitats can vary widely. A pH between 6.5 and 9.5 is acceptable for most species, with the optimum being between 7.2 and 8.5. Salinity should usually be below 5% of seawater (about 1.5 grams of sea salt per liter), but some species can tolerate much higher salinities, such as. D. magna, which can be found in up to 20% seawater.
Behavioral Ecology
The English name for Daphnia, waterflea, originates from the jumping-like behavior they exhibit while swimming. This behavior stems from the beating of the large antennae, which they use to direct themselves through the water. The rapid downbeat produces a quick upward movement, whereas the relatively high density of the animals creates a sinking. Motionless Daphnia sink rapidly to the ground.
A second well-known behavior of Daphnia is diel vertical migration, in which they migrate toward upper levels of the water body during nighttime and then back downward during the early morning and daytime. This behavior probably developed as a predator avoidance strategy. During daylight, the Daphnia hide from fish that hunt visually by moving to darker depths, whereas during nighttime, they take advantage of the richer food (planktonic algae) in the well-illuminated upper water levels. Inverse diel vertical migration has been described as a strategy to escape other predators that migrate themselves.
Part of the behavioral repertoire of Daphnia that is key to diel vertical migration is phototaxis. Clones of D. magna vary strongly in their phototactic behavior. Phototactic-positive genotypes spend much time in the upper water level, whereas phototactic-negative genotypes spend most of their time close to the bottom sediments. Phototactic behavior is also influenced by the presence of fish. If Daphnia sense that fish are present in the water, they behave more phototactically negative than they would otherwise.
Daphnia are also known to migrate toward or away from the banks of ponds or lakes. Again, the most likely explanation for this behavior is predator avoidance.
Certain species of Daphnia, e.g., D. magna, can be observed clinging occasionally to plants or other substrate. Furthermore, they may browse over the surface substrates to pick up small particles. This behavior is more apparent when food is limited and seems to enrich the diet. The stirring movement of the Daphnia brings small particles into suspension, which are then ingested by filter feeding.
Evolutionary Genetics
Following the pioneering work of P.D.N. Hebert, the population genetics of Daphnia have been intensively studied around the world. This study was facilitated by allozyme electrophoresis (more recently also microsatellite loci), which revealed a fair degree of enzyme polymorphism in many populations and opened the door for many studies on migration and gene flow (e.g., population divergence, isolation by distance, F-statistics), hybridization (many Daphnia species tend to form hybrids), inbreeding (small pools may be colonized by a few clones that subsequently inbreed), and clonal selection (clone frequency changes across the summer season have been frequently observed). It also helped explain phylogenetic relationships among species (later refined with DNA data).
Because of their clonal reproduction, Daphnia present a superb tool for quantitative genetic studies, which can enhance our understanding of their evolutionary ecology. Within- and between-clone comparisons can demonstrate genetic variation for various traits within and between populations, thus helping to reconstruct the evolutionary history of a population. For nearly every trait that has been investigated, genetic variation has been reported. Examples include age and size at maturity, size at birth, aging, reaction norms for life history traits, vertical migration, phototactic behavior, fish escape behavior, production of defense spines and helmets, resistance against parasites, immune response, competitive ability, Malthusian growth rate, carrying capacity under stable conditions, and many more. Furthermore, complex matrices of genetic covariances among traits have shown that, in a first approximation, most traits share some covariance with other traits, indicating that an evolutionary change of many traits is constrained by the evolution of other traits.
For those Daphnia species that have been karyologically investigated, between 20 and 24 chromosomes have been counted (2N) (Zaffagnini 1987).
With the announcement that the genome sequence of D. pulex will be produced in 2005, the genetic study of Daphnia is about to enter a new phase. With Daphnia, one of the first organisms with a well-known ecology will be sequenced, which is a milestone in the field of ecological genetics. Other molecular tools have been developed in parallel, not only for D. pulex but also for D. magna (e.g., microarrays, expressed sequence tags (ESTs)).
Population Dynamics
Daphnia populations vary strongly in density throughout the growing season. They typically go through pronounced cycles, with densities varying by more than seven orders of magnitude within a single season. A number of studies have refuted the belief that cycles are largely influenced by abiotic conditions; now it is commonly believed that abiotic conditions play a role only in limiting the growing season, not as a factor shaping the population dynamics during the growing season. In many populations, density peaks are observed two or even three times per year, and it is not uncommon for populations to disappear entirely during unfavorable seasons. Although it is difficult to make generalizations about Daphnia population dynamics, a few points are apparent.
In most habitats, Daphnia have low density or completely disappear during part of the year, usually the cold or the dry season. Recruitment in the following growing season is from resting eggs and/or from surviving females. There is rapid population increase in the early season (exponential growth), with doubling times of a few days (down to 3 days at temperatures of 20ºC and above). Growth is eventually slowed down by density-dependent competition, usually because of food shortage; however, predators may contribute as well. During this part of the season, parasites seem to play little role in affecting population numbers. The peak in Daphnia density usually follows a peak in algae density and may be followed by the clear-water phase in which the Daphnia effectively remove most of the phytoplankton from the water. The resulting food shortage leads to a rapid decline in Daphnia density. In large eutrophic lakes in temperate regions, phytoplankton and Daphnia may go through two density cycles (a spring and a summer peak), whereas in nutrient-poor lakes, only one peak may occur in mid-season. In small water bodies such as rock pools and vernal pools, the dynamics may look very different, depending on the expected length of the growing season. In pools with a very short growing season (e.g., Arctic and desert pools), Daphnia populations may produce resting eggs after only one or two generations, which curtails the exponential growth phase early. In longer-lasting small pools, populations may go through several population cycles within one season. Factors that increase population growth rate (e.g., eutrophication) or that amplify the response to high population density (e.g., sexual reproduction) increase the likelihood of more cycles occurring.
Daphnia parasites are most commonly observed after the first peak in population density. However, it is not clear whether parasites influence the population dynamics of Daphnia in natural populations. Parasites are an attractive ecological force for Daphnia population regulation because transmission is often density dependent, and they are found in nearly every Daphnia population investigated thus far. Experimental epidemiology with various zooplankton parasites has shown that parasites not only suppress host density but also may bring host populations to extinction (Ebert et al. 2000). Thus, it seems likely that the dynamics of natural Daphnia populations are influenced by parasites as well.
References
- Ebert D , Lipsitch M , Mangin KL . The effect of parasites on host population density and extinction: Experimental epidemiology with Daphnia and six microparasites. Am Nat. 2000;156:459–477. [PubMed: 29587512]
- Freyer G . Functional morphology and the adaptive radiation of the Daphniidae (Branchiopoda: Anomopoda). Philos Trans R Soc. 1991;331:1–99.
- Kästner. 1993. Lehrbuch der speziellen Zoologie, Band 1, Teil 4. Heidelberg, Germany, Elsevier GmbH, Spektrum Akademischer Verlag.
- Kükenthal W, Matthes E. 1944. Leitfaden für das Zoologische Praktikum. Jena, Germany, Gustac Fischer Verlag.
- Lampert W, Sommer U. 1999. Limnoökologie, 2. edition. Stuttgart, Thieme Verlag.
- Little TJ , O'Connor B , Colegrave N , Watt K , Read AF . Maternal transfer of strain-specific immunity in an invertebrate. Curr Biol. 2003;13:489–492. [PubMed: 12646131]
- Metchnikoff E . Ueber eine Sprosspilzkrankheit der Daphniden. Beitrag zur Lehre über den Kampf der Phagocyten gegen Krankheitserreger. Virchows Arch Pathol Anat Physiol. 1884;96:177–193.
- Mucklow PT , Ebert D . The physiology of immunity in the water flea Daphnia magna: Environmental and genetic aspects of phenoloxidase activity. Physiol Biochem Zool. 2003;76:836–842. [PubMed: 14988798]
- Peters RH, De Bernardi R. 1987. Daphnia. Memorie Dell 'Istituto Italiano di Idrobiologia. p. 502.
- Söderhall K . Editorial. Special issue. Invert Immun. 1999;23:263–266.
- Zaffagnini F . Reproduction in Daphnia. Mem Ist Ital Idrobiol. 1987;45:245–284.
- PubMedLinks to PubMed
- Introduction to Daphnia Biology - Ecology, Epidemiology, and Evolution of Parasi...Introduction to Daphnia Biology - Ecology, Epidemiology, and Evolution of Parasitism in Daphnia
Your browsing activity is empty.
Activity recording is turned off.
See more...