

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Varki A, Cummings RD, Esko JD, et al., editors. Essentials of Glycobiology. 2nd edition. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press; 2009.


Essentials of Glycobiology. 2nd edition.
Show details
Glycans are components of many biotherapeutic agents, ranging from natural products to molecules based on rational design to recombinant glycoproteins. The glycan components of these agents can be important determinants of their biological activity and therapeutic efficacy. This chapter provides a brief overview of some of the issues pertinent to glycan functions in therapeutics and a few examples of successful glycan or glycoconjugate-based drugs (Table 51.1).
TABLE 51.1
Examples of glycan-based drugs, their target diseases, and modes of action
GLYCANS AS COMPONENTS OF SMALL-MOLECULE DRUGS
Many well-known small-molecule drugs, such as antibiotics and anticancer therapeutic agents, are natural products that contain glycans as part of their essential structure and/or as a sugar side chain (i.e., a glycoside). Some examples of natural products that bear glycan side chains are shown in Figure 51.1. This is a well-established area of natural product and synthetic/biosynthetic chemistry and will not be reviewed in detail here. Glycans have also been modified to generate synthetic drugs. The most recent examples are the small-molecule inhibitors of influenza virus neuraminidase briefly discussed below (see also Chapter 50).
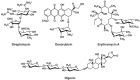
FIGURE 51.1
Examples of natural products that possess glycan components. Streptomycin and erythromycin A are antibiotics, doxorubicin is chemotherapeutic drug, and digoxin is used to treat cardiovascular disease.
SMALL-MOLECULE INHIBITORS OF INFLUENZA VIRUS NEURAMINIDASE
Influenza virus has two major surface proteins, termed hemagglutinin and neuraminidase (see Chapter 34). The hemagglutinin initiates infection by binding to cell-surface sialic acids. The neuraminidase assists virus release by cleaving sialic acids to prevent unwanted retention of newly synthesized virus on the cell surface. Neuraminidase may also function during the invasion phase by removing sialic acids on soluble mucins that would otherwise inhibit cell-surface binding. On the basis of the essential requirement for the neuraminidase during the viral life cycle, the crystal structure of the purified enzyme was studied and a rational drug design program was initiated. This approach resulted in the development of Zanamivir (Relenza™), in which the addition of a bulky guanidino side chain at the C-4 position of a previously known neuraminidase inhibitor, 2-deoxy-2,3-dehydro-N-acetyl-neuraminic acid, markedly increased the affinity for influenza neuraminidase, without having similar affects on host-cell neuraminidases (Figure 51.2; see also Chapter 50). Relenza blocks the life cycle of influenza virus by preventing infection and by interrupting the spread of the virus during the early phase of an infection. However, because it is not orally absorbable, the drug must be given as an inhaled medication to work at the mucosal sites of infection in the upper airway. An orally absorbable drug called Oseltamivir (Tamiflu™) was subsequently developed, and it can achieve the same effects. Both drugs are now in clinical use, either for treatment of influenza or for preventing spread of the disease.
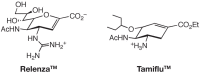
FIGURE 51.2
The synthetic influenza neuraminidase inhibitors Relenza™ and Tamiflu™.
The fear that the avian influenza virus (“bird flu”) might spread into the human population has resulted in stockpiling of Tamiflu. Although there is a public health rationale for this approach, it is difficult to know when and where the drug will be needed. In addition, drug-resistant strains of influenza are beginning to emerge, wherein the viral neuraminidase is no longer substantially inhibited. Thus, indiscriminate use of the drug might eventually, as in the case of antibiotics, negate its value when it is really needed. Moreover, widespread use may reveal off-target side effects, which may be more harmful to individuals for whom the risk of infection by influenza is low. Nevertheless, this is a classic example of how the study of the basic structural biology of a glycosidase and its glycan substrate resulted in one of the first effective drugs for a common and dangerous disease.
THERAPEUTIC GLYCOPROTEINS
A major proportion of biotherapeutic products are glycoproteins. These include erythropoietin and various other cytokines, antibodies, glycosyltransferases, and glycosidases, which together generate sales of billion of dollars worldwide. Therapeutic glycoproteins are typically produced as recombinant products in cell culture systems or, less commonly, in the milk of transgenic animals. Control of glycosylation is of major importance during the development of these drugs, because their glycan chains have marked effects on stability, activity, antigenicity, and pharmacodynamics in intact organisms. In most cases, glycosylation must be optimized to ensure prolonged circulatory half-life in the blood. Manipulation of glycans to promote targeting to specific tissues and cell types has also been a useful element of drug design.
Optimizing Glycans of Therapeutic Glycoproteins for Prolonged Serum Half-life
Erythropoietin is perhaps the most successful biotechnology product to date. It is a circulating cytokine that binds to the erythropoietin receptor, inducing proliferation and differentiation of erythroid progenitors in the bone marrow. Thus, it has much value in treating anemias caused by lack of erythropoietin (e.g., renal failure) or by bone marrow suppression (e.g., after chemotherapy). Natural and recombinant forms of erythropoietin carry three sialylated complex N-glycans and one sialylated O-glycan. Although in vitro the activity of deglycosylated erythropoietin is comparable to that of the fully glycosylated molecule, its activity in vivo is reduced by about 90%, because poorly glycosylated erythropoietin is rapidly cleared by filtration in the kidney. Undersialylated erythropoietin is also rapidly cleared by galactose receptors in hepatocytes and macrophages (see Chapter 31). These problems can be reduced by having fully sialylated chains and by increasing the amount of tetra-antennary branching. This, in turn, increases its activity in vivo nearly tenfold. Another approach is to deliberately add an N-glycosylation site to increase half-life and activity in vivo. Covalently linking polyethylene glycol to the protein also reduces clearance by the kidney.
Erythropoietin is somewhat unusual because it is small enough to be cleared by the kidney if it is underglycosylated. For most glycoprotein therapeutics, a more important consideration is minimizing clearance by galactose-binding hepatic receptors by ensuring full sialylation of glycans. Because glycans can have such dramatic effects on the properties of these drugs, it is important to ensure that glycosylation is controlled during their production, as well as to satisfy regulatory requirements for batch-to-batch product consistency. Changes in culture pH, the availability of precursors and nutrients, and the presence or absence of various growth factors and hormones can each affect the extent of glycosylation, the degree of branching, and the completeness of sialylation. Sialidases and other glycosidases that are either secreted or released by dead cells can also cause degradation of the previously intact product in the culture medium. Thus, during the development of a glycoprotein drug, considerable effort is devoted to defining appropriate conditions for reproducibly producing relatively homogeneous and complete glycosylation.
Impact of Glycosylation on Licensing and Patentability of Biotherapeutic Agents
Modern patenting of new therapeutics typically requires definition of the composition of matter in the claimed molecule. This is relatively straightforward for small molecules of defined structure and for nonglycosylated proteins. However, with regard to glycoproteins, especially those with multiple glycosylation sites, it is virtually impossible to obtain a single preparation that contains only a single glycoform. Thus, most biotherapeutic agents that are glycoproteins consist of a mixture of glycoforms. Licensing bodies such as the U.S. Food and Drug Administration have come to recognize this and will allow for a certain range of variation in glycoforms and the complexity of the mixture. However, the manufacturer and the agency must agree on the extent such variation is acceptable for a given drug formulation. Biopharmaceutical companies therefore spend considerable effort in assuring that their products fall within these defined ranges, once these are approved by licensing bodies. The inherent difficulty in reproducing complex glycoform mixtures also complicates efforts to make generic forms of recombinant glycoprotein drugs. Given the complexities of producing glycotherapeutic agents in mammalian cells, even the smallest changes in growth conditions (temperature, pH, media, etc.) can have significant effects on the range of glycoforms found in any given batch of the product. In fact, the licensing agencies also use consistency in glycoform composition as an indirect measure of the quality of process control in production. Differences in glycosylation can even have implications for the patentability of agents in which the underlying polypeptide remains constant. Marked differences in glycosylation have been used to define agents as being uniquely different. However, it is usually necessary to show that the differences in glycosylation being claimed also have a significant effect in changing the functionality of the drug in question. The associated pharmaceutical licensing and legal issues are rapidly evolving to keep pace with scientific advances in this area.
Significance of Nonhuman Glycosylation on Biotherapeutic Products
Certain types of nonhuman glycosylation, such as Galα1-3Gal (“α-Gal”) units and/or the sialic acid N-glycolylneuraminic acid (Neu5Gc), are found in terminal positions of glycans on some biotherapeutic glycoproteins. The addition of α-Gal residues can only occur if the therapeutic product is produced in a nonhuman cell line that expresses an α1-3Gal transferase not present in humans (see Chapter 13). The addition of Neu5Gc occurs when the nonhuman cell line used produces this sialic acid and/or because the animal-derived products used in the media provide a metabolic source of Neu5Gc, which can be taken up, processed, and made available for sialylation reactions (see Chapter 14). Other glycans unique to nonhuman expression systems may exist as well. Since humans have circulating antibodies against α-Gal termini, Neu5Gc, and presumably other nonhuman glycan determinants, a potential for antigen–antibody responses exists that could be deleterious and/or affect therapeutic efficiency. An optimal solution is to use cell lines that do not express α-Gal or Neu5Gc (e.g., CHO cells, which express no α-Gal and only low levels of Neu5Gc), to eliminate Neu5Gc from materials used in tissue culture, and to carefully analyze the glycans present in recombinant protein therapeutics. Further studies of the effects of these antigenic modifications on half-life and pharmacokinetics are needed.
GLYCOSYLATION ENGINEERING
There are limits as to how much of a biotherapeutic glycoprotein an animal cell line can produce. This becomes an issue especially when there is a need to produce very large amounts (e.g., if the glycoprotein needs to be given in high milligram quantities per day). In this situation, it would be beneficial to produce the glycoproteins in plants or yeast that are capable of much higher levels of production. However, it is necessary to eliminate risks arising from the nonhuman glycans of plant and fungal cells, which could cause excessively rapid clearance and/or antigenic reactions. Many plant and yeast glycans are immunogenic and elicit glycan-specific IgE and IgG antibodies in humans when delivered parenterally. To this end, there have been several efforts to add back a variety of mammalian genes into yeast and/or to eliminate genes that are producing nonhuman glycosylation. This has now been achieved using extensively engineered yeast strains that are capable of producing biantennary N-linked glycans with the human sialic acid N-acetylneuraminic acid (Neu5Ac). However, the productivity of such yeast strains has yet to be reported. Similar efforts to engineer yeast to make human-like O-glycans are under way, but other glycans such as glycosaminoglycans have not yet been addressed.
Plants and algae have also been used to engineer recombinant glycoproteins, but, as in yeast, the glycans produced by plants differ from those found in vertebrates. The antigenic differences that arise in recombinant glycoproteins produced in plants become less problematic if used for topical or oral administration, since humans are normally exposed to plant glycans in the diet. Another advantage is that the cost of production is much lower than in animal cell culture systems and animal sera are not needed. As in yeast, “humanizing plants” with respect to glycosylation may allow the production of nonimmunogenic glycoproteins. Chemical methods for synthesizing glycoproteins from scratch are also being explored as a means to obtain therapeutic agents with well-defined tailored glycans (see Chapter 49).
GLYCAN THERAPEUTIC APPROACHES TO METABOLIC DISEASES
Salvage versus De Novo Synthesis
All monosaccharides needed for cellular glycan synthesis can be obtained from glucose through metabolic interconversions (see Chapter 4). Alternatively, monosaccharides can be derived from the diet or salvaged from degraded glycans. The relative contributions of different sources can vary with the cell type. For instance, even though all mammalian cells use sialic acid, only some contain high amounts of UDP-GlcNAc epimerase/N-acetylmannosamine kinase (GNE), which is required for the de novo synthesis of CMP-sialic acid. But sialic acid salvage from degraded glycans is quite efficient, decreasing the demand on the de novo pathway. Similarly, galactose, fucose, mannose, N-acetylglucosamine, and N-acetylgalactosamine can come from the diet or be salvaged for glycan synthesis, whereas glucuronic acid, iduronic acid, and xylose cannot. All monosaccharides derived from the diet or degraded glycans can be catabolized for energy, and again, cells vary in their reliance on the different pathways.
The variable contributions of these pathways are important for therapy of some diseases. For instance, patients with congenital disorder of glycosylation type Ib (CDG-Ib), who are deficient in phosphomannose isomerase, benefit greatly from oral mannose supplementation to bypass the insufficient supply of glucose-derived mannose-6-phosphate. A few CDG-IIc patients have been treated with fucose to restore synthesis of sialyl Lewisx on leukocytes (see Chapter 42). Some patients with Crohn’s disease show clinical improvement with oral N-acetylglucosamine supplementation, but the mechanism is unknown. Mice deficient in GNE activity have kidney failure, but providing N-acetylmannosamine in the diet prevents this outcome. Clinical trials using N-acetylmannosamine to treat GNE-deficient patients with hereditary inclusion body myopathy type II (HIBM-II) are proposed, although it is unclear whether sialic acid deficiency is responsible for the human muscle pathology in HIBM-II.
Special Diets
Some monosaccharides and disaccharides can be toxic to humans who lack specific enzymes. For example, people who lack fructoaldolase (aldolase B) accumulate fructose-1-phosphate, which ultimately causes ATP depletion and disrupts glycogen metabolism. Prolonged fructose exposure in these people can be fatal, and fructose-limited diets are critical. Deficiencies in the ability to metabolize galactose (see Chapter 4) are mostly due to a severe reduction in galactose-1-phosphate uridyl transferase activity and cause galactosemia. Although these patients are asymptomatic at birth, ingesting milk leads to vomiting and diarrhea, cataracts, hepatomegaly, and even neonatal death. Low-galactose or galactose-free diets can prevent these life-threatening symptoms. However, even these diets do not prevent unexplained long-term complications, which include speech and learning disabilities and ovarian failure in almost 85% of females with galactosemia.
Infants hydrolyze lactose (Galβ1-4Glc) quite well, but the level of intestinal lactase can be much lower or absent in adults because of down-regulation of lactase gene expression. About two thirds of the human population has lactase nonpersistence, making milk products a dietary annoyance. This is because unabsorbed lactose provides an osmotic load and is metabolized by colonic bacteria, causing diarrhea, abdominal bloating and pain, flatulence, and nausea. Lactase persistence has evolved in certain pastoral populations from northwestern Europe, India, and Africa, allowing milk consumption in adult life. However, many adults either avoid lactose-containing foods or use lactase tablets to improve lactose digestion.
Substrate Reduction Therapy
The failure to turn over glycans by lysosomal degradation causes serious problems for patients with lysosomal storage disorders. Deficiencies in individual lysosomal enzymes lead to pathological accumulation of their substrates in inclusion bodies inside the cells (see Chapter 41). One approach to treating these disorders is to inhibit initial glycan synthesis, a strategy termed substrate reduction therapy (SRT). Reduced synthesis of the initial compound decreases the load on the impaired enzyme, and some patients show significant clinical improvement. One example of a drug used for SRT is N-butyldeoxynojirimycin (or N-butyl-DNJ) (Miglustat, Zavesca®), which has shown some efficacy for treating Gaucher’s disease (glucocerebrosidase deficiency).
Lysosomal Enzyme Replacement Therapy
Another approach for treating lysosomal storage disorders is enzyme replacement therapy. Unlike most therapeutic glycoproteins that interact with target receptors on the surface of cells, lysosomal enzymes developed for replacement therapy must be delivered intracellularly to lysosomes, their site of action. During the normal biosynthesis of lysosomal enzymes, their N-glycans become modified with mannose-6-phosphate (Man-6-P) residues, which target them to lysosomes using Man-6-P receptors (see Chapter 30). The challenge for enzyme replacement therapy is to get the enzymes targeted properly to lysosomes, where they can degrade accumulated substrate. A special case is enzyme replacement therapy for Gaucher’s disease. In this case, the enzyme has been successfully targeted to lysosomes of macrophages through their cell-surface mannose receptor (see Chapter 31). This approach required modification of the complex N-glycans of the enzyme isolated from placenta to expose terminal mannose residues, a process that involved digestion with various glycosidases (sialidase, galactosidase, and hexosaminidase). Nowadays, recombinant enzyme is produced in Lec1 mutant CHO cells (see Chapter 46), in which N-glycans with terminal mannose residues are present at all N-glycan sites.
The success of glucocerebrosidase treatment stimulated the development of lysosomal enzymes for treatment of other lysosomal storage diseases. Clinical trials have demonstrated clinical benefit in Fabry’s disease, mucopolysaccharidoses type I, II, and VI, and Pompe’s disease. However, the usefulness of replacement therapy is limited by the fact that injected enzymes do not have beneficial effects on all aspects of these diseases, and existing tissue damage is usually not reversible. Moreover, as Man-6-P receptors do not function in all cell types, N-glycan termini are needed to target all the affected cell types. In all instances, an additional challenge is to deliver enzymes efficiently to cells of the nervous system past the blood-brain barrier.
Chaperone Therapy
A third approach for treating lysosomal storage disorders takes advantage of the fact that some genetic defects lead to misfolding of the encoded enzyme in the endoplasmic reticulum (ER). There is evidence that low-molecular-weight molecules that are competitive inhibitors of some of these enzymes can act as “chaperones,” stabilizing the folded enzyme in the ER and effectively rescuing the mutation. The result is a higher steady-state concentration of active enzyme in the lysosome. Of course, the dose of such an inhibitor must be carefully adjusted to ensure that the inhibitory effects on enzyme function do not overshadow beneficial effects on folding. Fortunately, only a low level of enzyme restoration is needed to significantly reduce the accumulation of undigested glycan substrates, indicating that lysosomal hydrolases are normally present in large catalytic excess.
THERAPEUTIC APPLICATIONS OF GLYCOSAMINOGLYCANS
The use of purified glycans as therapeutics has received less attention than the development of glycoprotein-based treatments. Difficulties in establishing structure–activity relationships due to the large number of chiral centers and functional groups, undesirable pharmacokinetics of available formulations, poor oral absorption of the compounds, and low-affinity interactions with drug targets have limited their development. Some successful glycan drugs, such as the anticoagulant heparin, are given by injection, although efforts are under way to convert heparin into an orally absorbable form by complexing it with positively charged molecules. It may be possible to deliver other hydrophilic and/or negatively charged glycan drugs in this way to allow penetration of the intestinal barrier. Glycans are also sometimes attached to hydrophobic drugs to improve their solubility and alter their pharmacokinetics.
As discussed in Chapters 16 and 43, heparin is one of the most widely prescribed drugs today because of its anticoagulant activity, which is based on the binding and activation of antithrombin, a protease inhibitor of the coagulation cascade. Activation of antithrombin leads to rapid inhibition of thrombin and factor Xa, shutting down the production of fibrin clots. Heparin is produced by autodigestion of pig intestines, followed by graded fractionation of the products. Annual production is measured in metric tons, translating into a billion doses per year. Unfractionated heparin also binds to several plasma, platelet, and endothelial proteins, producing a variable anticoagulant response. Therefore, there has been much interest in producing heparin preparations with more predictable outcomes. Low-molecular-weight (LMW) heparins are derived by chemical or enzymatic cleavage of heparin to form smaller fragments. The pharmacological properties and the relative efficacy of the various LMW heparins appear generally superior to those of unfractionated heparin and there are fewer secondary complications. As the enzymes of heparin biosynthesis have now been cloned, it may be possible to produce recombinant heparins soon. There is also a synthetic heparin pentasaccharide that functions to bind antithrombin in exactly the same manner as heparin. This compound, known commercially as the drug Arixtra, is used to prevent deep-vein thrombosis and pulmonary embolism.
It is important to be able to neutralize heparin rapidly under some circumstances. The conventional approach uses the basic protein protamine, which binds to heparin, neutralizes its activity, and results in clearance of the complex by the kidney and liver. Recombinant heparinases are also under development as alternative neutralization agents. Heparin’s additional effects on blocking P- and L-selectin binding are discussed in Chapter 31. Heparin is also used to treat protein-losing enteropathy (PLE), likely working by competing for proinflammatory heparin-binding cytokines that trigger PLE in susceptible patients (see Chapter 43). It should be noted that the synthetic pentasaccharide Arixtra may not possess all of the nonanticoagulant activities of the natural products, such as selectin inhibition.
Hyaluronan (see Chapter 15) is a naturally occurring glycosaminoglycan that is extensively used in surgical applications. Because of its viscoelastic properties, hyaluronan has lubricating and cushioning properties that have made it useful for protecting the corneal endothelium during ocular surgery. Hyaluronan has antiadhesive properties and therefore might be useful in postsurgical wound healing as well. The mechanism of action is not well understood, but it may involve hyaluronan-binding proteins that mediate cell adhesion (see Chapter 15). Intra-articular injections of hyaluronan are used to treat knee and hip osteoarthritis. Several studies have shown modest improvement in patients treated with hyaluronan, but it is unclear if it is acting in a mechanical (as a viscosupplement) and/or a biological (via signaling pathways) manner. Hyaluronan is also injected as a tissue filler in cosmetic medicine.
“GLYCONUTRIENTS”
“Glyconutrient” is a term used by the nutritional supplement industry to describe some of their products. Numerous preparations are sold as nutritional supplements, often with wide-ranging claims concerning potential benefits. In most cases, these claims have not been substantiated through placebo-controlled, double-blind trials with defined, quantifiable outcomes. Much work is needed in this area to obtain insight into the potential role of dietary glycans on human health and to help consumers make wise decisions regarding their use.
As an example, mixtures of plant polysaccharides such as larchbark arabinogalactan, glucomannan, and others are often referred to “glyconutrients” that are claimed to contain “eight essential monosaccharides” needed for “cell communication.” Because all monosaccharides can be made from glucose (except in patients with rare genetic deficiencies; see Chapter 42), none of the other monosaccharides are actually known to be “essential.” Moreover, these polysaccharides are not degraded to available monosaccharides in the stomach or small intestine. Instead, anaerobic bacteria in the colon metabolize them and produce short-chain fatty acids. There are no peer-reviewed clinical studies supporting the efficacy of such “glyconutrients” for any disease or condition. Nevertheless, the following examples demonstrate how dietary glycans might have beneficial effects.
Glucosamine and Chondroitin Sulfate
Glucosamine (often mixed with chondroitin sulfate) has been promoted to relieve symptoms of osteoarthritis, which involves the age-dependent erosion of articular cartilage. Cartilage provides a cushion between the bones to minimize mechanical damage, and a net loss of cartilage occurs when the degradation rate exceeds the synthetic rate. A number of clinical trials report that glucosamine improves osteoarthritis symptoms, and some claim to restore partially the structure of the eroded cushion, in particular in the knees. Superficially, this would seem to make sense, because primary glycans of cartilage include hyaluronan (see Chapter 15) and chondroitin sulfate, both of which contain hexosamines within their structure (see Chapter 16). However, there are conflicting reports, and the outcome may depend on study design and the type and source of material (e.g., chloride vs. sulfate salts and the overall level of purity). Nevertheless, veterinarians have treated animals with glucosamine for over two decades with apparently positive results. Furthermore, double-blind, placebo-controlled studies in humans have shown a decreased rate of joint space narrowing. It should be noted that glucosamine might also alter UDP-GlcNAc and potentially UDP-GalNAc levels, thus affecting cellular responses involving any major class of glycans.
Positive effects of chondroitin sulfate on osteoarthritis are less well-documented. Indeed, how the acidic chondroitin sulfate polymer can be absorbed and delivered to its proposed site of action remains an open question. Further studies are needed to determine how dosage affects the circulating levels of these supplements, whether they are absorbed by the target tissue, and if they actually lead to changes in cartilage metabolism.
Xylitol and Sorbitol in Chewing Gum
Many studies suggest that chewing gum containing sugar alditols, specifically xylitol and sorbitol, can help control the development of dental caries. Mothers who chew xylitol-sweetened gum may even block transmission of caries-causing bacteria to their children. The benefit of these reduced sugars seems to be based on stimulation of salivary flow, but an antimicrobial effect is also possible. Xylitol also inhibits the expression and secretion of proinflammatory cytokines from macrophages and inhibits the growth of Porphyromonas gingivalis, one of the suspected causes of periodontal disease. Children who drank xylitol solutions also had a lower occurrence of otitis media.
Milk Oligosaccharides
Human milk contains about 70 g/liter of lactose and 5–10 g/liter of free oligosaccharides. More than 130 different glycan species have been identified with lactose at the reducing end, including poly-N-acetyllactosamine units. Some glycans are α2-3- and/or α2-6-sialylated and/or fucosylated in α1-2, α1-3, and/or α1-4 linkages. In contrast, bovine milk, the typical mainstay in human infant formulas, contains much smaller amounts of these glycans. These differences may account for some of the physiological advantages seen for breast-fed versus formula-fed infants. The glycans may also favor growth of a nonpathogenic bifidogenic microflora and/or block pathogen adhesion that causes infections and diarrhea. Surprisingly, a substantial number of human milk oligosaccharides remain almost undigested in the infant’s intestine and are excreted intact into the urine. Whether supplementing infant formula with specific, biologically active free glycans enhances infant health is unknown.
Chitosan
Chitosan (partially de-N-acetylated chitin, a polymer of β-linked N-acetylglucosamine residues) is a mixture of different-sized water-soluble polymers and has been reported to have hypocholesterolemic, antimicrobial, and weight-reduction effects. Combined analyses from various studies show a slight benefit in body weight reduction, but this benefit is less pronounced in more rigorous evaluations. The effects are likely to be of limited clinical significance.
GLYCANS AS VACCINE COMPONENTS
Microbial Vaccines
Vaccines consisting solely of glycan components typically elicit poor immunity, especially in infants. The primary limitation is that glycans are T-cell-independent antigens and therefore do not effectively stimulate T-helper-dependent activation and class switching of B-cell-mediated immunity. Conjugate vaccines with oligosaccharides coupled to carrier proteins have proven to be highly effective. For example, Haemophilus influenzae type b (Hib) causes an acute lower respiratory infection among young children. A conjugated form of an Hib-derived oligosaccharide coupled to a protein carrier is now routinely given to infants in developed countries and has resulted in a more than 95% decrease in incidence of infections in vaccinated populations. New vaccines are being developed using conjugated components of other bacterial capsular polysaccharides, and the studies appear promising.
Cancer Vaccines
Several carbohydrate-based vaccines are under development to treat cancer. Some of these vaccines are based on ganglioside immunogens present on certain types of cancer cells (e.g., gangliosides GM2 and GD2 in melanomas and globo H in breast cancer). In some cases, vaccines are composed of glycan-based haptens conjugated to a protein carrier. In one case, a breast cancer antigen sialyl-Tn (sialylα2-6GalNAcα-) is synthesized chemically and conjugated to a protein carrier (keyhole limpet hemocyanin). Sialyl-Tn is found on cancer mucins, and its expression correlates with progression to metastatic disease (see Chapter 44). A more direct strategy thus consists of attaching the sialyl-Tn units in their natural linkages to serine or threonine residues of the Muc-1 mucin polypeptide repeat. Although several cancer vaccine candidates are presently under investigation, they have yet to reach clinical utility.
BLOCKING GLYCAN RECOGNITION IN DISEASES
Blocking Infection
As discussed in Chapter 34, many microbes and toxins bind to mammalian tissues by recognizing specific glycan ligands. Thus, small soluble glycans or glycan mimetics can be used to block the initial attachment of microbes and toxins to cell surfaces (or block their release), and thus prevent or suppress infection. Because many of these organisms naturally gain access through the airways or gut, the glycan-based drugs can be delivered directly without the requirement of being distributed systemically. Examples of such applications currently under study include milk oligosaccharides that are believed to be natural antagonists of intestinal infection in infants (see above) and polymers that will block the binding of viruses such as influenza. Although backed by a strong scientific rationale and robust in vitro studies, such “antiadhesive” therapies have not yet found much practical application.
Inhibition of Selectin-mediated Leukocyte Trafficking
If specific glycan-protein interactions in vivo are responsible for selective cell–cell interactions and a resulting pathology, then infusion of small-molecule analogs or mimetics of the natural ligand could theoretically be useful. The best-studied example is the selectin-mediated recruitment of neutrophils (and other leukocytes) into sites of inflammation or ischemia/reperfusion injury, which involves specific selectin–glycan interactions occurring in the vascular system (see Chapter 31). Initial approaches focused on the use of sialyl Lewisx derivatives. However, the tetrasaccharide is a difficult synthetic target and has poor oral availability and a short serum half-life. To improve these shortcomings, attempts were made to design “glycomimetics,” which are compounds that preserve the essential functionality of the parent tetrasaccharide but eliminate unwanted polar functional groups and synthetically cumbersome glycan components. An example of the design of a monosaccharide glycomimetic starting from sialyl Lewisx is shown in Figure 51.3. First, the sialic acid residue was replaced with a charged glycolic acid group, the N-acetylglucosamine residue was then replaced with an ethylene glycol linker, and finally the galactose residue was replaced with a linker moiety. The resulting glycomimetic had E-selectin binding affinity comparable to sialyl Lewisx but with a simpler structure.
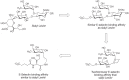
FIGURE 51.3
Glycomimetic E-selectin inhibitors based on sialyl Lewisx.
Simple monovalent compounds such as sialyl Lewisx and its glycomimetics have proven effective in various animal inflammatory models. This finding is somewhat surprising, because the binding constants for some of these monovalent ligands (with Kd values in the millimolar range) are much poorer than that for the physiological ligand. In fact, similar studies in humans did not show a clear benefit. Studies of the PSGL-1 (P-selectin glycoprotein ligand-1) glycosulfopeptide that has high affinity for P- and L-selectin are presently being pursued, as well as dendrimers consisting of multivalent forms of Lewis antigens. These conjugates have considerably longer serum half-lives than the monovalent glycans and glycomimetics. In addition, the inhibitory effects of heparin on P- and L-selectin are being considered. Finally, an alternate approach consists of finding specific inhibitors of the enzymes involved in forming endogenous ligand determinants in vivo. For example, the fucosyltransferase involved in sialyl Lewisx biosynthesis (FUT7) is an attractive target.
TRANSFUSION AND TRANSPLANTATION REJECTION BY ANTIGLYCAN ANTIBODIES
As discussed in Chapter 13, a variety of glycans, including the classical A and B blood group determinants, can act as barriers to blood transfusion and transplantation of organs. Rejection of mismatched blood or organs occurs because hosts have a high titer of preexisting antibodies against the glycan epitopes, presumably as a prior reaction to related structures found on bacteria or other microbes. In the case of the ABO blood groups, incompatibility is routinely managed by blood and tissue typing and finding an appropriate donor for the recipient. A successful strategy recently reported is to use bacterial enzymes in vitro to remove the A and B blood group determinants from A and B red cells, converting them into “universal donor” O red cells.
A related problem is found in xenotransplantation (i.e., the transplantation of organs between species) which is actively being pursued as a solution for the shortage of human organs for patients. The animal donors of preference are pigs, because many porcine organs resemble those of humans in size, physiology, and structure. However, unlike humans and certain other primates, pigs and most other mammals produce the terminal “α-Gal” epitope on glycoproteins and glycolipids. Because humans have naturally occurring high-titer antibodies in blood directed toward this epitope, this results in hyperacute rejection of porcine organ transplants, via reaction of the antibodies with endothelial cells of blood vessels. Attempts have been made to prevent this reaction, including blood filtration over glycan affinity columns to remove xenoreactive antibodies and blockade of the interaction by infusing soluble competing oligosaccharides. Transgenic pigs lacking the reactive epitope have also been produced, as have animals with an excess of complement-controlling proteins on their cell surfaces. Pig organs also have high levels of the nonhuman sialic acid (Neu5Gc), against which most humans have antibodies. Even if this problem is solved, there are other glycan and protein structural differences between humans and pigs that cause later stages of graft rejection, thus necessitating immunosuppression.
FURTHER READING
- Kunz C, Rudloff S, Baier W, Klein N, Strobel S. Oligosaccharides in human milk: Structural, functional, and metabolic aspects. Annu Rev Nutr. 2000;20:699–722. [PubMed: 10940350]
- Gomord V, Chamberlain P, Jefferis R, Faye L. Biopharmaceutical production in plants: Problems, solutions and opportunities. Trends Biotechnol. 2005;23:559–565. [PubMed: 16168504]
- Joshi L, Lopez LC. Bioprospecting in plants for engineered proteins. Curr Opin Plant Biol. 2005;8:223–226. [PubMed: 15753005]
- Mhurchu CN, Dunshea-Mooij C, Bennett D, Rodgers A. Effect of chitosan on weight loss in overweight and obese individuals: A systematic review of randomized controlled trials. Obes Rev. 2005;6:35–42. [PubMed: 15655037]
- Pastores GM, Barnett NL. Current and emerging therapies for the lysosomal storage disorders. Expert Opin. Emerg. Drugs. 2005;10:891–902. [PubMed: 16262569]
- Ly KA, Milgrom P, Rothen M. Xylitol, sweeteners, and dental caries [discussion 192–198] Pediatr Dent. 2006;28:154–163. [PubMed: 16708791]
- Beck M. New therapeutic options for lysosomal storage disorders: Enzyme replacement, small molecules and gene therapy. Hum Genet. 2007;121:1–22. [PubMed: 17089160]
- Brown JR, Crawford BE, Esko JD. Glycan antagonists and inhibitors: A fount for drug discovery. Crit Rev Biochem Mol Biol. 2007;42:481–515. [PubMed: 18066955]
- Butters TD. Pharmacotherapeutic strategies using small molecules for the treatment of glycolipid lysosomal storage disorders. Expert Opin Pharmacother. 2007;8:427–435. [PubMed: 17309337]
- Divine JG, Zazulak BT, Hewett TE. Viscosupplementation for knee osteoarthritis: A systematic review. Clin Orthop Relat Res. 2007;455:113–122. [PubMed: 17159579]
- Eklund EA, Bode L, Freeze HH. Diseases associated with carbohydrates/glycoconjugates. In: Kamerling JP, Boone GJ, Lee YC, editors. Comprehensive glycoscience. Vol. 4. Elsevier; New York: 2007. pp. 339–372.
- Hamilton SR, Gerngross TU. Glycosylation engineering in yeast: The advent of fully humanized yeast. Curr Opin Biotechnol. 2007;18:387–392. [PubMed: 17951046]
- Schultz BL, Laroy W, Callewaert N. Clinical laboratory testing in human medicine based on the detection of glycoconjugates. Curr Mol Med. 2007;7:397–416. [PubMed: 17584080]
- von Itzstein M. The war against influenza: Discovery and development of sialidase inhibitors. Nat Rev Drug Discov. 2007;6:967–974. [PubMed: 18049471]
- Schnaar RL, Freeze HH. A “glyconutrient sham. Glycobiology. 2008;18:652–657. [PubMed: 17855741]
- GLYCANS AS COMPONENTS OF SMALL-MOLECULE DRUGS
- SMALL-MOLECULE INHIBITORS OF INFLUENZA VIRUS NEURAMINIDASE
- THERAPEUTIC GLYCOPROTEINS
- GLYCOSYLATION ENGINEERING
- GLYCAN THERAPEUTIC APPROACHES TO METABOLIC DISEASES
- THERAPEUTIC APPLICATIONS OF GLYCOSAMINOGLYCANS
- “GLYCONUTRIENTS”
- GLYCANS AS VACCINE COMPONENTS
- BLOCKING GLYCAN RECOGNITION IN DISEASES
- TRANSFUSION AND TRANSPLANTATION REJECTION BY ANTIGLYCAN ANTIBODIES
- FURTHER READING
- Review Glycans in Biotechnology and the Pharmaceutical Industry.[Essentials of Glycobiology. 2022]Review Glycans in Biotechnology and the Pharmaceutical Industry.Seeberger PH, Freedberg DI, Cummings RD. Essentials of Glycobiology. 2022
- Review Glycoscience -- a new frontier in rational drug design.[Acta Pharm. 2006]Review Glycoscience -- a new frontier in rational drug design.Gornik O, Dumić J, Flogel M, Lauc G. Acta Pharm. 2006 Mar; 56(1):19-30.
- Review Glycans in Biotechnology and the Pharmaceutical Industry.[Essentials of Glycobiology. 2015]Review Glycans in Biotechnology and the Pharmaceutical Industry.Seeberger PH, Cummings RD. Essentials of Glycobiology. 2015
- Online nanoliquid chromatography-mass spectrometry and nanofluorescence detection for high-resolution quantitative N-glycan analysis.[Anal Biochem. 2012]Online nanoliquid chromatography-mass spectrometry and nanofluorescence detection for high-resolution quantitative N-glycan analysis.Kalay H, Ambrosini M, van Berkel PH, Parren PW, van Kooyk Y, García Vallejo JJ. Anal Biochem. 2012 Apr 1; 423(1):153-62. Epub 2012 Jan 28.
- Fluorescence-based solid-phase assays to study glycan-binding protein interactions with glycoconjugates.[Methods Enzymol. 2010]Fluorescence-based solid-phase assays to study glycan-binding protein interactions with glycoconjugates.Leppänen A, Cummings RD. Methods Enzymol. 2010; 478:241-64.
- Glycans in Biotechnology and the Pharmaceutical Industry - Essentials of Glycobi...Glycans in Biotechnology and the Pharmaceutical Industry - Essentials of Glycobiology
Your browsing activity is empty.
Activity recording is turned off.
See more...