By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Copyright © 2001, Sinauer Associates, Inc.
Bookshelf ID: NBK11148

An official website of the United States government

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Purves D, Augustine GJ, Fitzpatrick D, et al., editors. Neuroscience. 2nd edition. Sunderland (MA): Sinauer Associates; 2001.

Although receptor molecules that bind various tastants are found primarily on the apical microvilli of the taste cells, the transduction machinery involves ion channels on both the apical and basolateral membranes (Figure 15.12). Channels typically found in axonal membranes are located on the basolateral aspect of taste cells. These include voltage-gated Na+, K+, and Ca2+ channels that produce depolarizing potentials when taste cells interact with chemical stimuli. The resulting receptor potentials raise Ca2+ to levels sufficient for synaptic vesicle fusion and synaptic transmission, thus eliciting action potentials in the afferent axons. In general, the greater the tastant concentration, the greater the depolarization of the taste cell.
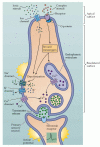
Transduction mechanisms in a generic taste cell. The apical and basolateral surfaces of the cell are separated by tight junctions. The apical surface contains both channels and G-protein-coupled receptors that are activated by chemical stimuli. The basolateral (more...)
The molecular identity of taste receptors has been examined in several experimental animals, including nonhuman primates. The “receptor” for salt (NaCl) is apparently an epithelial-type Na+ channel on the apical membrane of some taste cells (Figure 15.13). In general, the larger the NaCl concentration applied to the tongue, the larger the depolarization in the relevent taste cells. These Na+ channels are regulated by hormones involved in water and electrolyte balance (for example, antidiuretic hormone and aldosterone), which mediate Na+-specific appetite and intake. Protons (H+) can also diffuse through this channel, albeit more slowly than Na+; this fact may explain why the addition of acids like lemon juice to salty foods reduces their salty taste. Protons, which are primarily responsible for sour taste, also interact with distinct channels on the apical membranes of a subset of taste cells (see Figure 15.13). These cations activate proton-gated cation and Cl- channels (see Figure 15.12). Thus, several mechanisms underlie the reception and transduction of acidic stimuli (Figure 15.14).
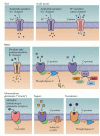
Examples of various channels and G-protein-coupled receptors that activate taste transduction in response to various compounds. When stimulated, each of these channels or receptors changes neurotransmitter release via either direct changes in depolarization (more...)

Different ways of encoding taste. (A) Response profiles of individual chorda tympani axons to four different stimuli (indicated by the four different colors). The numbers indicate individual axons. The responses reflect the net activity for 5 seconds (more...)
The transduction of sweet-tasting compounds involves the activation of G-protein-coupled receptors (GPCRs) on the apical surface of taste cells (see Figure 15.13). The particulars of the cascade depend on a number of factors, including the specifics of the stimulus. In the case of sweeteners such as the saccharides, activation of GPCRs depolarizes taste cells by activating adenylate cyclase, which in turn increases the cAMP concentration that will either directly or indirectly close basolateral K+ channels. Synthetic sweeteners, such as saccharine, activate different GPCRs that in turn activate phospholipase C (PLC) to produce IP3 and DAG. An increase in IP3 raises intracellular Ca2+ concentration, leading to transmitter release. An increase in DAG activates PKA, and PKA in turn phosphorylates and closes basolateral K+ channels, further contributing to this effect. Both of these pathways for the perception of sweetness can co-exist in the same taste cell.
There are many chemically distinct classes of bitter-tasting compounds (see Figure 15.13). Some of these are alkaloids, like quinine and caffeine; others are l-amino acids, urea, and even salts like MgSO4. Again, not all of these bitter tastants use the same receptor or transduction pathways. Indeed, about forty new receptors for bitter tastants have been recently cloned, and many are found in the same cells. Bitter- tasting organic compounds typically bind to GPCRs that activate gustducin (a G-protein found in taste cells homologous to transducin in photoreceptors), which in turn activates phosphodiesterase, thus lowering the cyclic nucleotide concentration and closing cyclic nucleotide-gated channels on the basolateral membranes of taste cells. Gustducin-knockout mice exhibit impaired responses to bitter compounds, suggesting that this second messenger is involved in the transduction of bitter tastes. Many bitter tastants bind directly to GCPRs that activate PLC and the production of IP3, leading to an increase in intracellular Ca2+. Given the wide variety of chemical structures that evoke a bitter taste, it is not surprising that a variety of transduction mechanisms are employed.
The taste of amino acids fall into both the sweet (d-amino acids) and bitter categories (l-isomers), and also uses a variety of transduction mechanisms. An exception is the amino acid l-glutamate (and its sodium salt), which elicits a quite different taste (see above). The effects of l-glutamate on taste cells involves both ionotropic receptors that activate ion channels, and unusual taste-specific metabotropic glutamate receptors (mGluR4) that are less sensitive to glutamate and that close ion channels through a cAMP-dependent pathway.
The overall picture that emerges from these admittedly complicated details is that taste cells have a variety of transduction mechanisms. In general, individual taste cells respond to several types of chemical stimuli. Nevertheless, taste cells also exhibit gustatory selectivity. Like olfactory cells, the lower the threshold concentration for detecting a single tastant, the greater the selectivity of the relevant taste cell. Finally, taste receptor mechanisms also adapt to the ongoing presence of a stimulus, although the mechanisms are not understood. If a chemical is left on the tongue for a sufficient time, it ceases to be perceived (consider saliva, for example). Thus, to obtain the full taste of foods, one must either frequently change the types of foods placed in the mouth or wait a sufficient time between helpings, facts that have long been appreciated by restauranteurs and gourmets.
By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Your browsing activity is empty.
Activity recording is turned off.
See more...